|
|
Навигация:
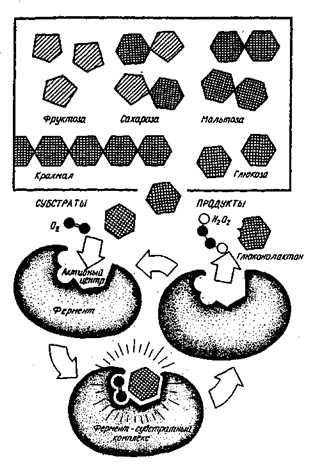
Биологические методы очистки сточных вод основываются на естественных процессах жизнедеятельности гетеротрофных микроорганизмов. Микроорганизмы, как известно, обладают целым рядом особых свойств, из которых следует выделить три основных, широко используемых для целей очистки: 2. Во-вторых, это свойство быстро размножаться. В среднем число бактериальных клеток удваивается через каждые 30 мин. По утверждению проф. Н.П. Блинова, если бы микроорганизмы могли беспрепятственно размножаться, то при наличии достаточного питания и соответствующих условий за 5 – 7 дней масса только одного вида микроорганизмов заполнила бы бассейны всех морей и океанов. Этого, однако, не происходит как из-за ограниченности источников питания, так и благодаря сложившемуся природному экологическому равновесию. 3. Способность образовывать колонии и скопления, которые сравнительно легко можно отделить от очищенной воды после завершения процессов изъятия содержавшихся в ней загрязнений. В живой микробиальной клетке непрерывно и одновременно протекают два процесса – распад молекул (катаболизм) и их синтез (анаболизм), составляющие в целом процесс обмена веществ – метаболизм. Иными словами, процессы деструкции потребляемых микроорганизмами органических соединений неразрывно связаны с процессами биосинтеза новых микробиальных клеток, различных промежуточных или конечных продуктов, на проведение которых расходуется энергия, получаемая микробиальной клеткой в результате потребления питательных веществ. Источником питания для гетеротрофных микроорганизмов являются углеводы, жиры, белки, спирты и т.д., которые могут расщепляться ими либо в аэробных, либо в анаэробных условиях. Значительная часть продуктов микробной трансформации может выделяться клеткой в окружающую среду или накапливаться в ней. Некоторые промежуточные продукты служат питательным резервом, который клетка использует после истощения основного питания. Весь цикл взаимоотношений клетки с окружающей средой в процессе изъятия из нее и трансформации питательных веществ определяется и регулируется соответствующими ферментами. Ферменты локализуются в Цитоплазме и в различных субструктурах, встроенных в мембрану клетки, выделяются на поверхность клетки или в окружающую среду. Общее содержание ферментов в клетке достигает 40-60% от общего содержания в ней белка, а содержание каждого из ферментов может составлять от 0,1 до 5% от содержания белка. При этом в клетках может находиться свыше 1000 видов ферментов, а каждую биохимическую реакцию, осуществляемую клеткой, могут катализировать 50-100 молекул соответствующего фермента. Часть ферментов представляют собой сложные белки (протеиды), содержащие кроме белковой части (апофермента) небелковую часть (кофер-мент). Во многих случаях коферментами являются витамины, иногда -комплексы, содержащие ионы металлов. Ферменты делятся на шесть классов по характеру реакций, катализирующих: окислительные и восстановительные процессы; перенос различных химических групп от одного субстрата к другому; гидролитическое расщепление химических связей субстратов; отщепление от субстрата химической группы или присоединение таковой; изменение в пределах субстрата; соединение молекул субстрата с использованием высокоэнергетических соединений. Поскольку микробиальная клетка потребляет только растворенные в воде органические вещества, то проникновение в клетку нерастворимых в воде веществ, таких, например, как крахмал, белки, целлюлоза и др. возможно лишь после их соответствующей подготовки, для чего клетка выпускает в окружающую жидкость необходимые ферменты для гидролитического их расщепления на более простые субъединицы. Коферменты определяют природу катализируемой реакции и по выполняемым функциям подразделяются на три группы: Механизм изъятия из раствора и последующей диссимиляции субстрата носит весьма сложный и многоступенчатый характер взаимосвязанных и последовательных биохимических реакций, определяемых типом питания и дыхания бактерий. Достаточно сказать, что многие аспекты этого механизма не совсем ясны до сих пор, несмотря на его практическое использование, как в области биотехнологии, так и в области биохимической очистки воды от органических примесей в широком спектре схем его технологического оформления. Наиболее ранняя модель процесса биохимического изъятия и окисления загрязнений основывалась на трех главных положениях: сорбционное изъятие и накопление изымаемого вещества на поверхности клетки; диффузионное перемещение через клеточную оболочку либо самого вещества, либо продуктов его гидролиза, либо гидрофобного комплекса образуемого гидрофильным проникающим веществом и белком-посредником; метаболическая трансформация поступивших внутрь клетки питательных веществ, обеспечивающая диффузионное проникновение вещества в клетку. В соответствии с этой моделью считалось, что процесс изъятия питательных веществ из воды начинается с их сорбции и накопления на поверхности клетки, для чего требуется постоянное перемешивание биомассы с субстратом, обеспечивающее благоприятные условия для “столкновения”^ клеток с молекулами субстрата. Механизм переноса вещества от поверхности клетки внутрь нее -эта модель объясняла либо присоединением проникающего вещества к специфическому белку-переносчику, являющемуся компонентом мембраны клетки, который после введения вещества внутрь клетки высвобождается и возвращается на ее поверхность для совершения нового “захвата” вещества и нового цикла переноса, либо непосредственным растворением этого вещества в веществе стенки и цитоплазматической мембраны, благодаря чему оно и диффундирует внутрь клетки. Процесс стабильного потребления вещества начинался лишь после некоторого “периода равновесия” вещества между раствором и клетками, объяснявшегося протеканием гидролиза и диффузионным перемещением вещества через клеточную оболочку до цитоплазматической мембраны, где сосредоточены различные ферменты. С началом метаболических превращений сорбционное равновесие нарушается, и концентрационный градиент обеспечивает непрерывность дальнейшего поступления субстрата в клетку. На третьем же этапе происходят все метаболические превращения субстрата частично в такие конечные продукты, как диоксид углерода, вода, сульфаты, нитраты (процесс окисления органических веществ), частично в новые микробиальные клетки (процесс синтеза биомассы), если процесс трансформации органических соединений происходит в аэробных условиях. Если же биохимическое окисление протекает в анаэробных условиях, то в его процессе могут образовываться различные промежуточные продукты (возможно целевого назначения), СН4, NH3, H2S и пр. и новые клетки. Эта модель, однако, не смогла объяснить некоторые кинетические особенности транспортных процессов переноса субстрата и, в частности, накопления субстрата в клетке против концентрационного градиента, являющегося наиболее частым результатом этих процессов и получившего название “активного” транспорта, в отличие от диффузионного переноса. Особенностью активных транспортных процессов является их стереоспе-Цифичность, когда близкие по химической структуре вещества конкурируют за общий переносчик, а не просто диффундируют в клетку под воздействием концентрационного градиента. В свете современных взглядов модель перемещения субстрата через клеточную мембрану предполагает наличие в ней гидрофильного “канала”, через который внутрь клетки могут проникать гидрофильные субстраты. Однако в отличие от вышеописанной модели здесь осуществляется стереоспецифическое перемещение, достигаемое, вероятно, за счет “эстафетной” передачи молекул субстрата от одной функциональной группы к другой. Субстрат при этом, как ключ, открывает соответствующий для его проникновения канал (модель трансмембранного канала). Вторая альтернативная модель может рассматриваться как комбинация первых двух с использованием их положительных свойств. В ней предполагается наличие гидрофобного мембранного переносчика, который путем последовательных конформационных изменений, вызываемых субстратом, проводит его с внешней на внутреннюю сторону мембраны (модель конформационной транслокации), где гидрофобный комплекс распадается. В данной интерпретации механизма транспорта субстрата через клеточную мембрану термин “переносчик” по-прежнему употребляется, хотя все чаще заменяется термином “пермеаза”, учитывающим генетическую основу его кодирования как мембранного компонента клетки для целей переноса вещества внутрь клетки. Установлено, что в состав мембранных транспортных систем часто входит более одного белкового посредника и между ними может существовать разделение функций. “Связующие” белки идентифицируют субстрат в среде, подводят и концентрируют его на внешней поверхности мембраны и передают его “истинному” переносчику, т.е. компоненту, осуществляющему перенос субстрата через мембрану. Так, выделены белки, участвующие в “узнавании”, связывании и транспорте ряда Сахаров, карбоновых кислот, аминокислот и неорганических ионов в клетки бактерий, грибов, животных. Превращение процесса переноса вещества в клетку в однонаправленный процесс “активного” транспорта, приводящий к повышению содержания питательных веществ в клетке против их концентрационного градиента в среде, требует от клетки определенных энергетических затрат. Поэтому процессы переноса субстрата из окружающей среды внутрь клетки сопряжены с протекающими внутри клетки процессами метаболического высвобождения заключенной в субстрате энергии. Энергия в процессе переноса субстрата расходуется на химическую модификацию либо субстрата, либо самого переносчика с тем, чтобы исключить или затруднить как взаимодействие субстрата с переносчиком, так и возврат субстрата диффузионным путем через мембрану обратно в раствор. Современные воззрения на процессы биохимического изъятия и окисления органических соединений основываются на двух кардинальных положениях теории ферментативной кинетики. Первое положение постулирует, что фермент и субстрат вступают во взаимодействие друг с другом, образуя фермент-субстратный комплекс, который в результате одной или нескольких трансформаций приводит к появлению продуктов, снижающих барьер активации катализируемой ферментом реакции за счёт её дробления на ряд промежуточных этапов, каждый из которых не встречает энергетических препятствий для своего осуществления. Второе положение констатирует то, что независимо от характера соединений и количества этапов в ходе ферментативной реакции, катализируемой ферментом, в конце процесса фермент выходит в неизменном виде и способен вступать во взаимодействие со следующей молекулой субстрата. Иными словами, уже на этапе изъятия субстрата клетка взаимодействует с субстратом с образованием относительно , непрочного соединения, называемого “фермент-субстратным комплексом”. Вышеуказанное хорошо иллюстрируется примером извлечения из раствора глюкозы различными микроорганизмами, содержащими фермент глюкозооксидазу в среде с молекулярным кислородом. Глюкозооксидаза образует фермент-субстратный комплекс – глюкоза – кислород – глюкозооксидаза, после распада которого образуются промежуточные продукты -глюконолактон и пероксид водорода, как это схематично показано на рис. 11.1. Образовавшийся в результате распада указанного комплекса глюконолактон подвергается гидролизу с образованием глюконовой кислоты. Одним из важнейших свойств ферментов является их способность синтезироваться при наличии и под воздействием определенного вещества. Другим не менее важным свойством является специфичность воздействия фермента как по отношению к катализируемой им реакции, так и по отношению к самому субстрату. Иногда фермент способен воздействовать на один единственный субстрат (абсолютная специфичность), но значительно чаще фермент воздействует на группу схожих по наличию в них определенных атомных группировок субстратов.
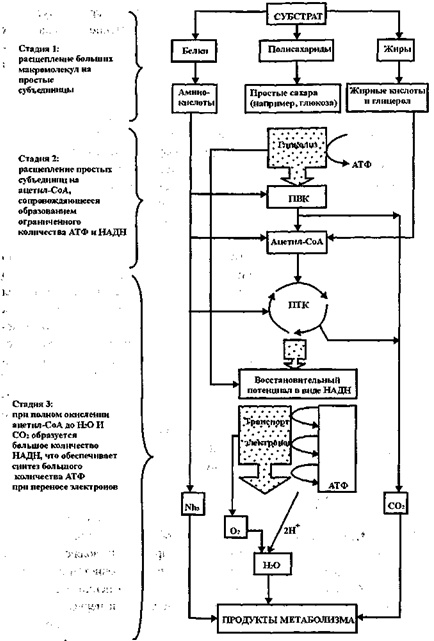
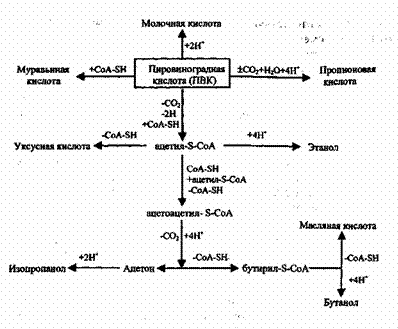
Рис. 11.1. Схема “узнавания” ферментом субстрата, образования фермент-субстратного комплекса и катализ Многим ферментам присуща стереохимическая специфичность, состоящая в том, что фермент воздействует на группу субстратов (а иногда на один), отличающихся от других особым расположением атомов в пространстве. Роль каждого фермента в процессе биохимического окисления органических веществ строго определенна: он катализирует либо окисление (т.е. присоединение кислорода или отщепление водорода), либо восстановление (т.е. присоединение водорода или отщепление кислорода) вполне определенных химических соединений. При дегидрировании тот или иной фермент может отщеплять лишь определенные атомы водорода, занимающие определенное пространственное положение в молекуле субстрата или промежуточного продукта. Сказанное относится и к ферментам, катализирующим другие метаболические процессы. Процессы биохимического окисления у гетеротрофных микроорганизмов делят на три группы в зависимости от того, что является конечным акцептором водородных атомов или электронов, отщепляемых от окисляемого субстрата. Если акцептором является кислород, то этот процесс называют клеточным дыханием или просто дыханием; если акцептор водорода -органическое вещество, то процесс окисления называют брожением; наконец, если акцептором водорода является неорганическое вещество типа нитратов, сульфатов и пр., то процесс называют анаэробным дыханием, или просто анаэробным. Наиболее полным является процесс аэробного окисления, т.к. его продукты – вещества, не способные к дальнейшему разложению в микро-биальной клетке и не содержащие запаса энергии, которая могла бы быть высвобождена обычными химическими реакциями. Главные из этих веществ, как уже отмечалось – диоксид углерода (С02) и вода (Н20). Хотя оба эти вещества содержат кислород, химический путь их образования в клетке может быть различным, поскольку диоксид углерода может получаться в результате биохимических процессов, протекающих в бескислородной среде под воздействием ферментов – декарбоксилаз, отщепляющих С02 от карбоксильной группы (СООН) кислоты. Вода же в результате жизнедеятельности клетки образуется исключительно путем соединения кислорода воздуха с водородом тех органических веществ, от которых он отщепляется в процессе их окисления. Аэробная диссимиляция субстрата – углеводов, белков, жиров -носит характер многостадийного процесса, включающего первоначальное расщепление сложного углеродсодержащего вещества на более простые субъединицы (к примеру полисахариды – в простые сахара; жиры – в жирные кислоты и глицерол; белки – в аминокислоты), подвергающиеся, в свою очередь, дальнейшей последовательной трансформации. При этом доступность субстрата окислению существенно зависит от строения углеродного скелета молекул (прямой, разветвленный, циклический) и степени окисления углеродных атомов. Наиболее легко доступными считаются сахара, особенно гексозы, за ними следуют многоатомные спирты (глицерин, маннит и др.) и карбоновые кислоты. Общий конечный путь, которым завершается аэробный обмен углеводов, жирных кислот, аминокислот, – цикл трикарбоновых кислот (ЦТК) или цикл Кребса, в который эти вещества вступают на том или ином этапе. Отмечается, что в условиях аэробного метаболизма около 90% потребляемого кислорода используется на дыхательный путь получения энергии клетками микроорганизмов. Брожение является процессом неполного расщепления органических веществ, преимущественно углеводов в условиях без кислорода, в результате которого образуются различные промежуточные частично окисленные продукты, такие как спирт, глицерин, муравьиная, молочная, про-пионовая кислоты, бутанол, ацетон, метан и др., что широко используется в биотехнологии для получения целевых продуктов. До 97% органического сУбстрата может превращаться в такие побочные продукты и метан. Ферментативное анаэробное расщепление белков и аминокислот называют гниением. Из-за малого выхода энергии при бродильном типе метаболизма, осуществляющие его микробиальные клетки должны потреблять большее количество субстрата (при меньшей глубине его расщепления), чем клетки, получающие энергию за счет дыхания, что объясняет более эффективный рост клеток в аэробных условиях по сравнению с анаэробными. Наибольшее количество энергии для своего функционирования клетка получает в результате окисления кислородом водорода, отщепляемого от окисляемого субстрата под действием ферментов-дегидрогеназ, которые по своему химическому действию делятся на никотинамидные (НАД) и флавиновые (ФАД). Никотинамидные дегидрогеназы первыми реагируют с субстратом, отщепляя от него два атома водорода и присоединяя их к коферменту. В результате этой реакции субстрат окисляется, а НАД восстанавливается до НАД‘Н2. Далее в реакцию вступает ФАД, перенося водород с никотинамидного кофермента на флавиновый, в результате чего НАД‘Н2 снова окисляется до НАД, а флавиновый – восстанавливается до ФАДН2. Далее через чрезвычайно важную группу окислительно-восстановительных ферментов-цитохромов – водород передается молекулярному кислороду, что и завершает процесс окисления с образованием окончательного продукта – воды. В этой реакции и высвобождается наибольшая часть заключенной в субстрате энергии. Весь процесс аэробного окисления может быть представлен схемой рис. 11.2. Высвобождающаяся в процессе микробиального окисления вещества энергия аккумулируется клеткой с помощью макроэргических соединений. Универсальным накопителем энергии в живых клетках является аденозинтрифосфорная кислота – АТФ (хотя имеются и другие магроэнерги). Эта реакция фосфорилирования, как видно из (11.9) нуждается в энергии, источником которой в данном случае является окисление. Поэтому фосфорилирование АДФ тесно сопряжено с окислением, в связи чем этот процесс называют окислительным фосфорилированием. В процессе окислительного фосфорилирования при окислении, например, одной молекулы глюкозы образуется 38 молекул АТФ, тогда как в стадии гликолиза -только 2. При этом следует отметить, что стадия гликолиза протекает совершенно одинаково и в аэробных, и в анаэробных условиях, т.е. до образования пировиноградной кислоты (ПВК), и на его протекание затрачиваются 2 из 4 образующихся молекул АТФ. Пути дальнейшей трансформации ПВК в аэробных и в анаэробных условиях расходятся. Аэробная трансформация глюкозы может быть представлена следующей схемой: Окисление НАДН2 в системе переноса электронов дает ЗАТФ на В условиях анаэробного превращения углеводов первым этапом является фосфорилирование глюкозы, осуществляемое с помощью АТФ под воздействием фермента гексокиназы, т.е. Окислительное фосфорилирование может осуществляться и под воздействием фермента, синтезирующего АТФ на уровне субстрата. Однако, такое образование макроэргических связей носит весьма ограниченный характер, и в присутствии кислорода клетки синтезируют большую часть содержащейся в них АТФ через систему переноса электронов. Аккумуляция высвобождающейся в процессе диссимиляции вещества в аэробных или анаэробных условиях с помощью макроэргических соединений (и прежде всего АТФ) позволяет устранить несоответствие между равномерностью процессов высвобождения химической энергии из субстрата и неравномерностью процессов ее расходования, неизбежной в реальных условиях существования клетки. Упрощенно весь процесс распада органических веществ в ходе аэробных превращений может быть представлен схемой, приведенной на рис. 11.3. Схема же анаэробных превращений ПВК после стадии гликолиза представлена на рис. 11.4. Исследованиями установлено, что зачастую тип метаболизма зависит не столько от наличия кислорода в среде, сколько от концентрации субстрата. Это указывает на то, что в зависимости от конкретных условий функционирования биомассы в среде могут одновременно протекать как аэробные, так и анаэробные процессы трансформации органических соединений, интенсивность которых также будет зависеть от концентрации и субстрата и кислорода. Здесь следует отметить, что в промышленной биотехнологии для получения различных продуктов микробиального происхождения (кормовых или пекарских дрожжей, различных органических кислот, спиртов, витаминов, лекарственных препаратов) используются чистые культуры, т.е. микроорганизмы одного вида зачастую селекционируемые, со строгим поддержанием видового состава, соответствующих условий питания, температуры, активной реакции среды и пр., исключающих появление и развитие других видов микроорганизмов, что могло бы привести к отклонению качества получаемого продукта от установленных стандартов. При очистке же сточных вод, содержащих смесь разнообразных по химическому составу загрязнений, которые иногда даже весьма трудно идентифицировать аналитическими методами, биомасса, осуществляющая очистку, также представляет собой смесь, а точнее, сообщество различных видов микроорганизмов и простейших со сложными между ними отношениями. Как видовой, так и количественный состав биомассы очистных сооружений будет зависеть от конкретного метода биологической очистки и условий его реализации. По расчетам некоторых специалистов, при концентрации растворенных органических загрязнений, оцениваемых показателем БПКП0Лн, до 1000 мг/л наиболее выгодно применение аэробных методов очистки. При концентрациях БПКПОЛн от 1000 до 5000 мг/л экономические показатели аэробных и анаэробных методов будут практически одинаковыми. При концентрациях же свыше 5000 мг/л более целесообразным будет применение анаэробных методов. Однако, при этом следует принимать во внимание не только концентрацию загрязнений, но и расходы сточных вод, а также тот факт, что анаэробные методы приводят к образованию таких конечных продуктов, как метан, аммиак, сероводород и др. и не позволяют получить качество очищенной воды, сопоставимое с качеством очистки аэробными методами. Поэтому при высоких концентрациях загрязнений применяется сочетание анаэробных методов на первой ступени (или первых ступенях) очистки и аэробных методов на последней ступени очистки. Следует подчеркнуть, что бытовые и городские сточные воды, в отличие от производственных, не содержат концентраций загрязнений, оправдывающих применение анаэробных методов, и потому эти методы очистки в данной главе не рассматриваются.
Рис. 11.3. Упрощенная схема трехстадийного распада молекул питательных веществ (Б. Альберте и др. 1986)
Рис. 11.4. Превращение пировиноградной кислоты анаэробными микроорганизмами в различные продукты Похожие статьи: Навигация: Статьи по теме:
Главная → Справочник → Статьи → Блог → Форум |
|
|
|
|
Информационный сайт о строительных материалах и технологиях. Контакты: Никита Королёв - © 2008-2014 |
© Все права защищены.
Копирование материалов невозможно. |
|